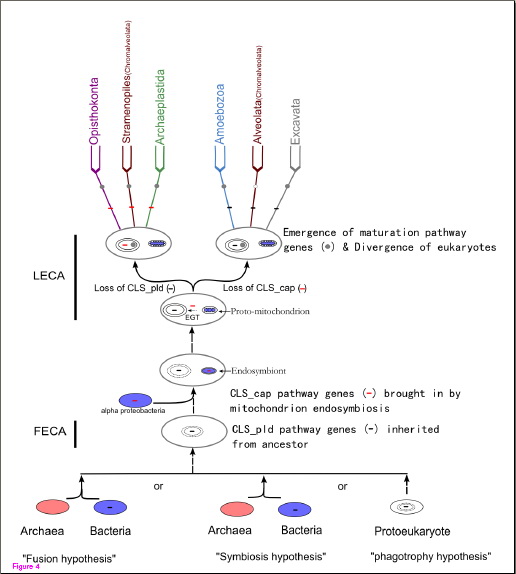
图:心磷脂合成和成熟途径在整个生物界中的演化历史
心磷脂(cardiolipin),同时是真核细胞线粒体内膜和原核细胞细菌膜上的重要结构和功能成分。该事实被认为是支持线粒体是由细菌内共生起源而来的重要证据之一。然而,一般认为心磷脂的生物合成在细菌和真核生物中分别是由具有不同结构域的两种不同类型的心磷脂合成酶(CLS),即CLS_pld和CLS_cap,来催化完成的。那么,这就产生了一个重要问题:为什么真核细胞线粒体心磷脂的合成途径不是由细菌内共生直接带入却出现不同?从原核生物进化成真核生物的过程中,真核生物的心磷脂合成途径究竟有着怎样的起源?随着真核生物的分化,该途径又经历了怎样的进化呢?
中国科学院昆明动物研究所真核细胞进化基因组学研究组田海峰博士在导师文建凡研究员的指导下,首先对心磷脂合成及其成熟酶在整个生物三大部类(Domain)中的分布进行了系统地调查。很有趣地发现:1)尽管大部分真细菌中存在的是CLS_pld途径,但是在包括变形菌和放线菌等一部分真细菌中已经出现了过去被认为是真核型的CLS_cap途径;2)尽管所有多细胞生物和部分单细胞真核生物普遍具有CLS_cap途径,但在很多单细胞的原生生物中却只具有过去被认为是原核型的CLS_pld途径;3)心磷脂成熟酶为真核生物所特有而不存在于原核生物中。其次,分子系统分析结果显示,真核生物所具有的CLS_cap途径与α蛋白菌的CLS_cap途径的亲缘关系最为接近;部分单细胞原生生物中发现的CLS_pld途径虽然也与细菌中的CLS_pld途径关系密切但不能确定来源于哪一类特定的细菌;心磷脂成熟酶均是在真核生物起源后通过别的基因的duplication等途径而进化出来的。据以上研究事实,他们勾勒出了心磷脂合成和成熟途径在整个生物界中的演化历史脉络图:真核生物的最早共同祖先(FECA)从原核祖先中获得了CLS_pld途径;之后,在真核生物最近共同祖先(LECA)中,内共生的线粒体又带来了CLS_cap途径;此时,不同的原生生物出现了不同的选择,其中选择了CLS_cap的成为了有机会进化成多细胞真核生物的原生生物类群。心磷脂成熟途径出现在真核生物产生之后,而且,该途径在不同生物中的多样性可能是适应性进化的结果;而“无线粒体”原生生物(如蓝氏贾第虫)中缺少心磷脂合成与成熟途径,很可能是因为二次丢失造成的。这些研究结果对加深我们认识真核生物代谢途径的进化具有重要意义。该研究成果近期已在“BMC Evol. Biol.” (2012, 12:32)上发表。
该研究得到了科技部973,国家自然科学基金委和中国科学院研究项目的资助。
